|
|
実績・論文など
発表論文(2008年の主な論文)
(1) Depletion of nucleophosmin leads to distortion of nucleolar and nuclear structures in HeLa cells, Amin MA.; Matsunaga, S.; Uchiyama S.; Fukui, K. Biochem. J. 2008 415(3), 345-351.
(2) Live cell imaging reveals plant Aurora kinase has dual roles during mitosis, Kurihara, D.; Matsunaga, S.; Uchiyama, S.; Fukui, K. Plant Cell Physiol. 2008 49(8), 1256-1291.
(3) Nucleophosmin is required for chromosome congression, proper mitotic spindle formation, and kinetochore-microtubule attachment in HeLa cells, Amin MA.; Matsunaga, S.; Uchiyama S; Fukui K. FEBS Lett. 2008, 582(27), 3839-44.
(4) Genome Structure of the Legume, Lotus japonicus, Sato, S., Nakamura, Y., Kaneko, T., Asamizu, E., Kato, T., Nakao, M., Sasamoto, S., Watanabe, A., Ono, A., Kawashima, K., Fujishiro, T., Katoh, M., Kohara, M., Kishida, Y., Minami, C., Nakayama, S., Nakazaki, N., Shimizu, Y., Shinpo, S., Takahashi, C., Wada, T., Yamada, M., Ohmido, N., Hayashi, M., Fukui, K., Baba, T., Nakamichi, T., Mori, H. and Tabata, S. DNA Res. 2008 15(4), 2030-40
(5) Visualization of mitotic HeLa cells by advanced polarized light microscopy, Morimoto, A.; Matsunaga, S.; Kurihara, D.; Fukui, K. Micron, 2008 39(5), 635-638.
(6) Improvement of bioactive beads-mediated transformation by concomitant application of electroporation, Murakawa, T.; Kajiyama, S.; Fukui, K. Plant Biotechnol. 2008 105, 77-80.
(7) Protection of isolated chromosomes using calcium-alginate microbeads (bioactive beads) toward chemical and physical stresses, Murakawa, T.; Hayashihara, K.; Ito, K.; Doi, T.; Kajiyama, S.; Fukui, K. Chromosome Sci., 2008 10, 83-87.
(8) Preparation Methods of Human Metaphase Chromosomes for their Proteome Analysis, Fukui, K.; Takata, H.; Uchiyama, S. Methods Mol. Biol., 2008 432, 149-60.
(9) Improvement of transformation efficiecy in bioactive beads-mediated gene transfer using DNA-lipofectin complex as entrapped genetic material, Murakawa, T.; Kajiyama, S.; Ikeuchi, T.; Kawakami, S; Fukui, K. J. Biosci. Bioeng. 2008, 105(1), 77-80.
(10) In vivo manipulation of fluorescently-labelled organelles in living cells by multi-photon excitation, Watanabe, W.; Matsunaga, S.; Higashi, T.; Fukui, K.; Itoh, K. J. Biomed. Opt. 2008 13, 31213-31221.
(11) The Arabidopsis SDG4 contributes to the regulation of pollen tube growth by methylation of histone H3 lysines 4 and 36 in mature pollen, Cartagena, J.A.; Matsunaga, S.; Seki, M.; Kurihara, D.; Yokoyama, M.; Shinozaki, K.; Fujimoto, S.; Azumi, Y.; Uchiyama, S.; Fukui, K. Dev. Biol. 2008, 315(2), 355-368.
(12) Anti-Peptide Antibodies for Examining the Conformation, Molecular Assembly and Localization of an Intracellular Protein, Ribosomal Protein S6, In vivo., Nakagawa, M.; Ohmido, N.; Ishikawa, K., Uchiyama, S.; Fukui, K. ;Azuma, T. J. Biochem. 2008 143(3), 325-332.
染色体高次構造の解明とその利用を目指して
染色体高次構造の謎を解く
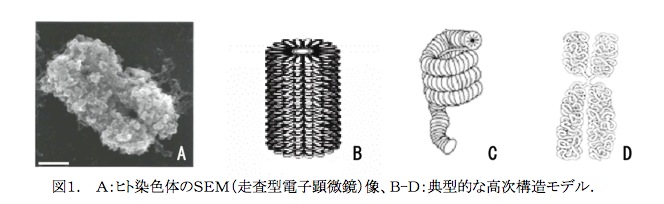
19世紀に遺伝情報の可視的担体である染色体(図1A)が発見されて以来、その生物学的重要性については既に多くの論文で証明されてきた。その為、染色体は現在の分子細胞生物学の最も重要な研究対象となっている。しかしながら、多くの仮説が図1B、1C、1D.に示すように提案されているものの、21世紀なる今日においてもその高次構造については明らかにされていない。我々の研究対象はまさにこの染色体の高次構造であり、考えうる種々の方法を用いてこの3世紀にわたる謎を解こうと努力している。
観察手段の革新
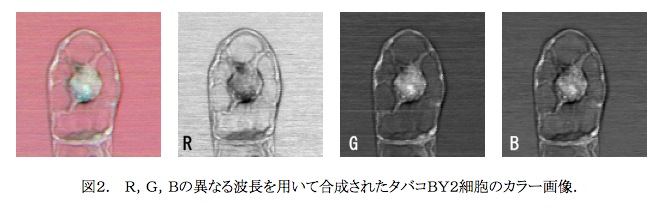
染色体の画像解析システムCHIAS1-4を自ら開発し、定量的かつ正確な染色体画像解析を進めてきた。特に態細胞分裂前中期の植物小型染色体に典型的に認められる凝縮型(Condesation pattern,CP)を画像解析することにより、イネ・アラビドプシスをはじめとする多くの植物染色体が世界で初めて識別・同定された。現在はさらに生命先端工学専攻伊東研究室と共同し、全く新しい光学顕微鏡を開発に成功している。それは, 4つの光波を混合して用いるものであり、誘導パラメトリック発光(Stimulated Parametoric Emission,SPE)顕微鏡と命名された。本顕微鏡は生物試料を ①生きたまま、②無染色にて、③3次元的に観察する、ことを可能とするものであり、その発展型である誘導ラマン散乱顕微鏡(SRS)の開発とともに、生体資料観察手段に大きな革新をもたらすものである。SPE顕微鏡で最初にとられた、無染色植物培養生細胞の疑似カラー画像を図2.に示す。
染色体構成タンパク質の謎を解く
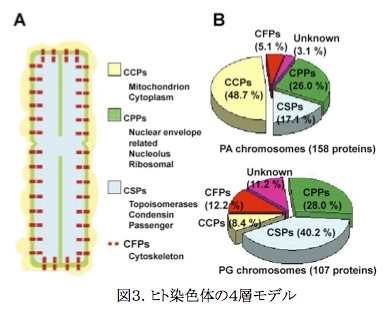 染色体は構成成分は2分子のDNAと塩基性のヒストンタンパク質および非ヒストンタンパク質である。しかしながら非ヒストンタンパク質にどのようなものがどの程度含まれているかは知られておらず、染色体構成タンパク質の全体像を把握することは染色体の高次構造を知る上で必要不可欠のステップであった。われわれは、そこで染色体構成タンパク質のプロテオーム解析を世界に先駆けて行い、200種以上の染色体タンパク質をその含量とともに同定した。 またこれらタンパク質の性質・機能を検討することにより、これらを、4つのグループに分別することが可能であることを明らかにした。
染色体は構成成分は2分子のDNAと塩基性のヒストンタンパク質および非ヒストンタンパク質である。しかしながら非ヒストンタンパク質にどのようなものがどの程度含まれているかは知られておらず、染色体構成タンパク質の全体像を把握することは染色体の高次構造を知る上で必要不可欠のステップであった。われわれは、そこで染色体構成タンパク質のプロテオーム解析を世界に先駆けて行い、200種以上の染色体タンパク質をその含量とともに同定した。 またこれらタンパク質の性質・機能を検討することにより、これらを、4つのグループに分別することが可能であることを明らかにした。
その上で、これら4グループのタンパク質が染色体上の特徴的な位置を占めること、およびその機能と深く関連していることを見出した。 4つのグループはそれぞれChromo-some coating protein (CCP)、Chromosome peripheral protein (CPP)、Chromosome structural protein (CSP)、Chromosome fibrous protein (CFP)と命名された(図3B)。 この発見により、ヒト染色体は、構成タンパク質の面から見ると図3A.に示す4層構造をとることが明らかになり、ヒト染色体の「4層モデル」が提案された。
染色体タンパク質の動的解析
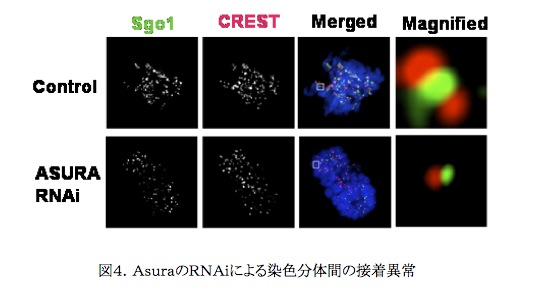
プロテオーム解析により同定された染色体タンパク質はそれらの抗体を作製しつつ、その細胞周期を通じた局在解析および機能解析の対象とされた。RNAi(RNA干渉)はタンパク質の機能解析に大きな威力を発揮した。RNAiによる解析を通じて、それぞれのタンパク質が細胞周期の進行、染色体動態、核形態、党に従来知られていなかった多くの機能を有することが明らかとなってきた。例えば、アスラと命名したタンパク質(Prohibitin 2,PHB2) は、図4.に示すように細胞分裂時の姉妹染色分体間の接着に関係していることが明らかとなった。もともとアスラは多機能を有することが知られていたが、細胞分裂時におけるこうした機能については今回が初めての報告となった。